

Elektronenförderkettenkomponente, Sequenz, Inhibitoren
- 2226
- 418
- Rieke Scheer
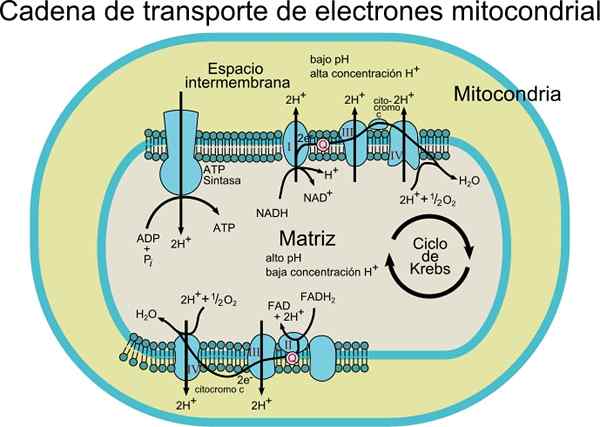
Der Elektronenförderkette Es besteht aus einer Reihe von Protein- und Coenzymmolekülen in einer Membran. Wie der Name schon sagt, ist es für den Transport von Elektronen aus dem NADH- oder FADH2 -Coenzyme zum endgültigen Empfänger, der O2 (Molekular Sauerstoff) ist, verantwortlich.
In diesem Transportprozess ist die Energie, die durch die Übertragung von Elektronen aus Coenzymen auf molekularer Sauerstoff durch Redox -Zentren mit Protein übertragen wird, mit der Energieproduktion (ATP) verbunden. Diese Energie wird dank des Protonengradienten erhalten, der in der mitochondrialen Innenmembran erzeugt wird.
Dieses Transportsystem besteht aus verschiedenen Komponenten, die in mindestens zwei Oxidationszuständen gefunden werden können. Jeder von ihnen wird effektiv reduziert und während der Bewegung von Elektronen von NADH oder FADH2 nach O2 reoxyd.
NAD+ und FAD -Coenzyme sind in Fettsäuroxidationswegen und des Zitronensäurzyklus infolge der Oxidation verschiedener Substrate reduziert. Anschließend sind diese Coenzyme in der elektronischen Transportkette oxidiert.
So dass das elektronische Transportsystem aus einer Abfolge von Oxid-Reduktions-Reaktionen besteht, die miteinander verbunden sind.
[TOC]
Kettenkomponenten
Abhängig von der Art des Organismus können 3 bis 6 Komponenten beobachtet werden, die die Elektronenförderkette bilden. Der Elektronentransportprozess und die ATP -Synthese durch oxidative Phosphorylierung sind Prozesse, die in einer Membran auftreten.
Bei prokaryotischen Zellen (aerobe Bakterien) treten diese Prozesse mit der Plasmamembran auf. In Eukaryot -Zellen tritt sie in der Mitochondrienmembran auf, sodass die Elektronentransportkomponenten im inneren Teil der Membran gefunden werden.
https: // giphy.com/gifs/htksbbedquk9z7co5e
Elektronen werden allmählich über vier Komplexe übertragen, aus denen die elektronische Transportkette besteht.
Jeder Komplex hat mehrere Proteinkomponenten, die mit prothetischen Gruppen (nicht -Aminosäurekomponenten von konjugierten Proteinen) assoziiert sind, die es ermöglichen, dass ihre Reduktionspotentiale zunehmen.
Darüber hinaus besteht dieses Transportsystem aus verschiedenen molekularen Spezies wie Flavoprotein; Coenzym, das auch Ubiquinona (Coq oder UQ) bezeichnete; Verschiedene Cytochrome wie Cytochrom B, C, C1, A und A3; Proteine mit Fe-S und Proteingruppen, die mit Cu verbunden sind. Diese Moleküle sind mit der Membran verbunden, mit Ausnahme von Cytochrom C.
Komplex i
https: // giphy.com/gifs/rlf1rl9gdc6356sj6w
Der I-Komplex namens Nadh Coenzym Quinona oxidortaduase oder Nadh-Dehydrogenase besteht aus rund 45 Polypeptidketten und enthält ein mononukleotid-Flavinmolekül (FMN) und acht bis neun Fe-S-Gruppen Fe-S. Wie der Name schon sagt, überträgt dieser Komplex ein Elektronenpaar vom Nadh Coenzym auf Coq.
Die Funktion des NADH -Dehydrogenase -Komplexes beginnt mit der Vereinigung von NADH zu diesem Komplex an der Seite der inneren mitochondrialen Membranmatrix. Die Elektronen werden dann von NADH zum FMN transportiert. Anschließend passieren Elektronen vom reduzierten Flavin (FMNH2) zu Fe-S-Proteinen.
Das FMNH2 fungiert als eine Art Brücke zwischen dem NADH und den Fe-S-Proteinen, da letztere nur ein einzelnes Elektron übertragen können, während das Nadh Coenzym zwei überträgt, so dass die Flavinas diese Übertragung eines einzelnen Elektrons dank seiner Redox durchführen des Semi -Kynones.
Kann Ihnen dienen: Monocytopoese: Stufen, Merkmale, RegulierungSchließlich werden die Elektronen von den Fe-S-Gruppen auf das Coenzym Q übertragen, einem mobilen Elektronenförderer mit einem Isopreneid-Schwanz, der es hydrophob macht, das die Mitte der Mitochondrienmembran überqueren zu.
Komplex II
![]()
Der II -Komplex, der besser als Succinat -Dehydrogenase bekannt ist, ist ein integrales Protein der mitochondrialen Innenmembran und ein Enzym, das in den Zitronensäurezyklus interveniert.
Dieser Komplex besteht aus zwei hydrophilen Untereinheiten und zwei hydrophoben HEMO-B.
Im Zitronensäurezyklus (Krebs oder Tricarbonsäurezyklus) wird Succinat durch Succinat -Dehydrogenase in Fumarat umgewandelt, wodurch das Coenzym FAD auf FADH2 reduziert wird. Aus diesem letzten Coenzym werden Elektronen in die Fe-S-Zentren übertragen, die sie wiederum in den Coq übertragen.
Während der Reaktionen dieses Elektronentransfers ist das Standard -Redoxpotential sehr niedrig, was verhindert, dass die notwendige freie Energie freigesetzt wird, um ATP zu synthetisieren.
Dies bedeutet, dass Komplex II der einzige Komplex in der Elektronentransportkette ist, der keine Energie für die ATP -Synthese bereitstellen kann. Dieser Komplex ist jedoch der Schlüssel im Prozess, da er Elektronen von FADH2 auf den Rest der Kette überträgt.
Komplex III
https: // giphy.com/gifs/cjg0raiuizjds2hkyu
Der III -Komplex, Cytochromkomplex BC1 oder CoQ Cytochrom C -Reduktase, überträgt die Elektronen aus dem reduzierten Coenzym zum Cytochrom C. Diese Übertragung erfolgt über eine einzelne Redoxstraße, die als Q -Zyklus bezeichnet wird.
Dieser Komplex besteht aus einem Protein mit Fe-S und drei verschiedenen Cytochromen, in denen das in der HEMO-Gruppe befindliche Eisenatom zyklisch zwischen den reduzierten Zuständen (Fe2+) und oxidiertem (Fe3+) variiert.
Cytochrome sind Hämoproteine des elektronischen Transports, die eine Redoxaktivität aufweisen. Sie sind in allen Organismen vorhanden, mit Ausnahme einiger obligatorischer Anaerobes.
Diese Proteine haben Hämogruppen, die zwischen zwei Oxidationszuständen abwechseln (Fe2+ und Fe3+). Cytochrom C ist ein mobiler Elektronentransporter, der schwach mit der internen Membran von Mitochondrien assoziiert ist.
Die in diesem Komplex gefundenen Cytochrome sind Cytochrome B, C und A, die 3 sind aktive Redoxproteine mit Gruppen. Wir haben unterschiedliche Eigenschaften, die ihre Oxidationszustände zwischen Fe2+ und Fe3 abwechseln+.
Cytochrom C ist ein peripheres Membranprotein, das mit Cytochrom C1 und mit dem IV -Komplex als „Shuttle“ des Elektrons fungiert.
IV -Komplex
Cytochrom C und O2 sind die endgültigen Rezeptoren von Elektronen, die aus der Oxidation des organischen Materials abgeleitet sind, so. Dies akzeptiert Elektronen aus Cytochrom C und überträgt sie auf O2 -Reduktion.
Die komplexe Funktion besteht darin, die Oxidationen eines Elektrons der vier aufeinanderfolgenden Moleküle von reduziertem Cytochrom C zu katalysieren, dh gleichzeitig vier Elektronen eines O2 -Moleküls reduziert und schließlich zwei H2O -Moleküle produzieren.
Kann Ihnen dienen: EpithelbeschichtungsgewebeElektronentransportsequenz
Die Elektronen werden dank des Coenzyms Q von den Komplexen I und II auf den Komplex III übertragen, und von dort übergeben sie durch Cytochrom C zum IV -Komplex C. Wenn die Elektronen diese vier Komplexe durchlaufen, nimmt das Reduktionspotential zu und freisetzt Energie, die dann für die ATP -Synthese verwendet wird.
Insgesamt verursacht die Übertragung einiger Elektronen die Translokation von 10 Protonen durch die Membran; Vier in den Komplexen I und IV und zwei des Komplexes III.
Nadh Descidrogenasa
Dieses Enzym katalysiert die Oxidation des Nadh Coenzyms durch das Coenzym, das. Die Elektronen bewegen sich von der NADH zum FMN, das am hydrophilen Schwanz des Komplexes i angebracht ist. Fe-S-Gruppierungen übertragen auf Elektronen gleichzeitig. Diese Fe-S-Gruppen reduzieren sich auf den COQ, der in die Membran eingebettet ist, in Ubiquinol (reduzierter COQ).
Während der Übertragung von Elektronen auf den COQ werden vier Protonen wiederum über die interne Membran auf den intermembranalen Raum übertragen. Der Mechanismus, durch den diese Protonen transloziert sind.
Der Elektronenübertragungsprozess in diesem Schritt fördert freie Energie, insbesondere -16,6 kcal/mol.
CoQ-Citocromo C-Reduktase und Q-Zyklus
Das Coenzym, das durch Cytochrom C in einer von diesem Coenzym katalysierten Reaktion oxidiert wird. Die Oxidation des Ubiquinols (reduzierten COQ) tritt in einer bestimmten Stelle des Komplexes (QO oder Oxidationsstelle) in der Mitochondrienmembran auf und überträgt zwei Elektronen, eines mit den Fe-S-Gruppen auf das Protein und eine in die HEMO-Gruppen auf das Protein.
Im Q -Zyklus erzeugt die Oxidation von COQ Semi -Archinon, wobei die Elektronen in die HEMO B1- und BH -Gruppen übertragen werden. Während dieser Elektronentransfer tritt ein zweiter COQ in der QO -Stelle oxidiert und wiederholt den Zyklus.
Dieser Zyklus bewirkt die Übertragung von zwei Elektronen und die Translokation von vier Protonen in den intermembranalen Raum, wobei die Freisetzung von -10,64 kcal/mol freier Energie freisetzt.
Citocroma -C -Oxidase
Dieses Enzym (Komplex IV) katalysiert die Oxidation von Cytochrom C (reduziert) durch O2, was der endgültige Elektronenakzeptor ist. Diese Übertragung erzeugt ein H2O -Molekül für jedes Elektronenpaar, das zusätzlich zur Translokation von Protonen über die Membran übertragen wird.
Die Elektronen bewegen sich von eins zu eins, vom reduzierten Cytochrom C bis zu einem Paar Cua -Ionen, dann übergeben sie zu einer Hämo -A -Gruppe und erreichen schließlich das binukleare Zentrum des Komplexes, der Cub- und Hemo -A3 -Ionen enthält, wobei die Übertragung von vier Übertragungen enthält Elektronen treten Sauerstoff auf.
Im IV -Komplex transferieren die Elemente die Elektronen von eins nacheinander.
Die in diesem Stadium freigesetzte Energie entspricht -32 kcal/mol. Der während des Übertragungsprozesses erzeugte elektrochemische Gradienten und die Energieänderungen (ΔE) entsprechen in jeder Stufe der freien Energie, die für die Produktion eines ATP -Moleküls erforderlich ist.
Kann Ihnen dienen: MembranfaltungstheorieGroße Dehydrogenase
Wie bereits erwähnt, hat dieser Komplex die einzige aber wichtige Funktion der Einführung der FADH2 -Elektronen vom Zitronensäurezyklus in die Elektronentransportkette.
Dieses Enzym katalysiert die Oxidation des FADH2 -Coenzyms durch das Coenzym q (oxidiert). Im Zitronensäurezyklus werden zwei Elektronen und zwei Protonen in die Modeerscheinung übertragen. Anschließend überträgt der FADH2 diese Elektronen bis COQ über die Fe-S-Zentren des Komplexes.
Schließlich werden die Elektronen aus dem COQ in den Komplex III übertragen, folgt den oben beschriebenen Schritten.
Die Kettenkomplexe sind unabhängig
Die vier Komplexe, aus denen die elektronische Transportkette besteht.
Die I- und II -Komplexe bewegen sich in der Membran, indem sie ihre Elektronen auf CoQ übertragen, die sich auch in der Membran ausbreiten und sie in Komplex III übertragen, von wo aus Elektronen bis zum Cytochrom C übergehen, das auch in der Membran mobil ist und Elektronen im IV -Komplex abliegt.
Inhibitoren der elektronischen Transportkette
Auf der elektronischen Transportkette wirken einige spezifische Inhibitoren in ihrem Prozess. Rothenon ist ein häufig verwendetes Insektizid, das stöchiometrisch mit dem Komplex I verbunden ist, wodurch die Reduktion von COQ vermieden wird.
Einige Barbiturate-Arzneimittel wie Piericidin und Amytal hemmen den Komplex I und stören die Übertragung von Elektronen aus den Fe-S-Gruppen auf den COQ.
In Komplex II wirken einige Verbindungen wie das Öltrifluoraceton und Malonat als Wettbewerbsinhibitoren mit Succinat, um ihre Oxidation zu vermeiden und die Elektronenübertragung auf die Modeerscheinung zu übertragen.
Einige Antibiotika wie Myxotiazol und Stigmatellin binden an die Q-Stelle des COQ, wobei die Übertragung von Elektronen aus dem Coenzym q zu den Fe-S-Zentren der Proteine hemmt.
Cyanid, Azida (N3-), Schwefelsäure und Kohlenmonoxid hemmen den IV-Komplex. Diese Verbindungen binden an Hämogruppen und vermeiden die Übertragung von Elektronen in das Binuklearzentrum des Komplexes oder Sauerstoffs (O2) (O2).
Wenn die Elektronenförderkette gehemmt wird, wird die Energieerzeugung aufgrund der oxidativen Phosphorylierung gestoppt, was schwerwiegende Schäden und sogar den Tod des Organismus verursacht.
Verweise
- Alberts, geb., Bray, d., Hopkin, k., Johnson, a., Lewis, J., Raff, m., Roberts, k. & Walter, p. (2004). Essentielle Zellbiologie. New York: Garlandwissenschaft. 2. Ausgabe.
- Cooper, g. M., Hausman, r. UND. & Wright, n. (2010). Die Zelle. (pp. 397-402). Ed. Marbán.
- Devlin, t. M. (1992). Lehrbuch der Biochemie: mit klinischen Korrelationen. John Wiley & Sons, Inc.
- Garrett, r. H., & Grisham, C. M. (2008). Biochemie. Ed. Thomson Brooks/Cole.
- Rawn, j. D. (1989). Biochemie (Nr. 577.1 roh). Ed. Interamerikanische McGraw-Hill
- Voet, d., & Voet, j. G. (2006). Biochemie. Ed. Pan -American Medical.
- « Elektrische Anguila -Eigenschaften, Lebensraum, Reproduktion
- Biozoos -Eigenschaften, Morphologie, Fortpflanzung, Ernährung »