

Cardiolipin -Struktur, Synthese, Funktionen
- 2065
- 149
- Lewis Holzner
Der Cardiolipin, Er ist auch als Defathidylglycerin bekannt, er ist ein Lipid der Familie der Glycepholipide und der Gruppe von Polyglyphospholipiden. Es kommt in der mitochondrialen Membran eukaryotischer Organismen, in der Plasmamembran vieler Bakterien und auch in einigen Bögen vor.
Es wurde 1942 von Pangborn aus der Analyse der Membranlipide des Herzgewebes eines Rinderes entdeckt. Die Struktur wurde 1956 vorgeschlagen und die chemische Synthese fand etwa 10 Jahre später statt.
Cardiolipin -Struktur (Quelle: Edgar181 [Public Domain] über Wikimedia Commons) Einige Autoren sind der Ansicht, dass ihre Anwesenheit auf ATP-produzierende Membranen beschränkt ist, wie es bei Mitochondrien in Eukaryoten, Plasmamembranen in Bakterien und Wasserstoffosomen (Typ-Mitocondria-Organellen) in bestimmten Protisten der Fall ist.
Die Tatsache, dass Cardiolipin in den Mitochondrien und in der Plasmamembran von Bakterien liegt die Zelle und umgekehrt.
Sein Biosyntheseweg bei Tieren wurde zwischen 1970 und 1972 beschrieben, und anschließend wurde gezeigt, dass es dieselbe Route ist, der in Pflanzen, Hefen, Pilzen und Wirbellosen auftritt. Es ist kein sehr reichlich vorhandenes Lipid, aber die Zellen benötigen, um richtig zu funktionieren.
Die Bedeutung dieses Phospholipids für Mitochondrien und daher für den Zellstoffwechsel ist deutlich, wenn die mit ihm verbundene fehlerhafte Funktionen von Stoffwechselstrecken eine menschliche Pathologie erzeugen, die als Barth -Syndrom bekannt ist (kardiooskelettisches Myopathie).
[TOC]
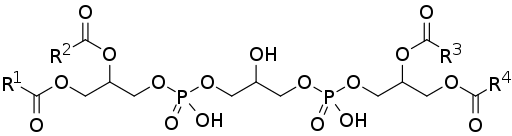
Struktur
Cardiolipin oder Defospathidylglycerin besteht aus zwei Phosphatidsäure (leichtere Phospholipid), die durch ein Glycerinmolekül miteinander verbunden sind.
Phosphatidsäure, eine der üblichen Vermittler auf den Biosynthesewegen anderer Phospholipide, besteht aus einem 3-phosphat-Glycerolmolekül, zu dem zwei Ketten von Fettsäuren in den Positionen der Karbonen 1 und 2 verärgert werden, so dass es auch als 1,2 als 1,2 bekannt ist -Diacylglycerin 3-phosphat.
Kann Ihnen dienen: Sekundärstruktur von Proteinen: EigenschaftenDaher besteht Cardiolipin aus drei Glycerinmolekülen: einem zentralen Glycerol zusammen mit einer Phosphatgruppe in Kohlenstoff 1, einer anderen Phosphatgruppe in Kohlenstoff 3 und einer Hydroxylgruppe in Kohlenstoff 2; und zwei "laterale" Glycerines.
Die beiden "lateralen" Glycerolmoleküle binden durch "Glycerolbrücken" durch ihre Kohlenstoffe in Position 3 an das zentrale Molekül. In Kohlenstoffen der Positionen 1 und 2 haben zwei Ketten von Fettsäuren und variable Sättigung verestert.
Cardiolipin ist ein Lipid, das je nach Anwesenheit oder Abwesenheit von trennen Kationen Bicapas bilden kann oder nicht. Dies hängt mit der Tatsache zusammen, dass es sich um ein symmetrisches Molekül handelt, das es für Membranen wichtig ist, die für Energieübertragungsprozesse verantwortlich sind.
Wie die anderen Lipide der Gruppe der Polyglyphospholipide hat Cardiolipin mehrere Hydroxylgruppen, die für die Vereinigung von Fettsäuren dienen können. Daher hat es mehrere Positionssteroisomere.
Seine Fettsäuren
Unterschiedliche Studien haben festgestellt, dass Fettsäuren, die an die lateralen Glycerinmoleküle von Cardiolipin gebunden sind.
Dann können solche Fettsäuren zwischen 14 und 22 Längenkohlen und zwischen 0 und 6 Doppelbindungen haben. Dies und die Tatsache, dass Cardiolipin vier gebundene Fettmoleküle hat, impliziert, dass es mehrere Variablen und Kombinationen dieses Phospholipids geben kann.
Synthese
Die Biosynthese von Cardiolipin beginnt, wie zu erwarten war. Dieser Prozess tritt in den Mitochondrien von Eukaryoten und in der Plasmamembran von Bakterien auf.
Kann Ihnen dienen: Cyclasa Adenilate: Eigenschaften, Typen, FunktionenSynthese in Eukaryoten
Nach der Bildung reagiert Phosphatidsäure mit einem hohen Energiemolekül analog zu ATP: CTP. Dann ist ein Vermittler mit hoher Energie als Phosphatidyl-CMP bekannt. Die aktivierte Phosphatidil -Gruppe wird in der C1 -Position des zentralen Glycerolmoleküls auf die Hydroxylgruppe übertragen, das als Skelett dient.
Dieser Prozess ergibt sich. Dies empfängt eine andere Phosphatidylgruppe, die von einem anderen Phosphatidyl-CMP-Molekül aktiviert ist, einer Reaktion.
Das Synthase -Cardiolipin -Enzym liegt in der inneren mitochondrialen Membran und scheint zumindest in Hefen einen großen Komplex zu bilden. Sein Gen wird in großen Mengen in reichen Stoffen in Mitochondrien wie Herz, Leber und Skelettmuskel von Wirbeltieren exprimiert.
Die Regulation seiner Aktivität hängt weitgehend von denselben Transkriptionsfaktoren und endokrinen Faktoren ab, die die mitochondriale Biogenese modulieren.
Einmal in der inneren mitochondrialen Membran synthetisiert, muss Cardiolipin in die externe mitochondriale Membran transloziert werden, sodass eine Reihe topologischer Prozesse in der Membran angegeben sind und andere strukturelle Elemente derselben angepasst sind.
Synthese in Prokaryoten
Der Cardiolipingehalt in Bakterien kann sehr variabel sein und hängt hauptsächlich vom physiologischen Zustand der Zellen ab: In der exponentiellen Wachstumsphase ist er normalerweise weniger häufig und häufiger vorhanden, wenn es reduziert wird (zum Beispiel in der stationären Phase), bei der Verringerung der stationären Phase vorhanden ist.
Sein Biosyntheseweg kann von verschiedenen stressigen Reizen wie Energiedefizit oder osmotischer Stress erschossen werden.
Bis zur Bildung von Phosphatidylglycerin ist der Prozess in Eukaryoten und Prokaryoten äquivalent, aber in den Prokaryoten erhält das Phosphatidylglycerin durch Transcessterifizierung eine Phosphatidylgruppe aus einem anderen Phosphatidylglycerolmolekül. Diese Reaktion wird durch eine Enzymphospholipase D katalysiert, die auch als Cardiolipin -Synthase bekannt ist.
Es kann Ihnen dienen: SGLT (Natriumglucose-Transportproteine)Diese Reaktion ist als "Transfatidilation" -Reaktion (aus Englisch "bekanntTransposphatidylierung "), Wo einer der Phosphatidylglycerinolen als Spender der Phosphatidil -Gruppe und der andere als Akzeptor fungiert.
Funktionen
Die physikalischen Eigenschaften von Cardiolipinmolekülen ermöglichen anscheinend bestimmte Wechselwirkungen, die wichtige Funktionen in der strukturellen Organisation der Membranen ausführen, in denen sie gefunden werden.
Zu diesen Funktionen gehört die Unterscheidung einiger Membrandomänen, Wechselwirkung oder "Kreuzung" mit Transmarket -Proteinen oder ihren Subdomänen unter anderem.
Dank seiner physikalisch -chemischen Eigenschaften wird Cardiolipin als Lipid anerkannt, das keine Bápara bildet, deren Funktion jedoch die Stabilisierung sein kann und "von Transmembranproteinen in Lipidbikapas untergebracht" sein kann.
Insbesondere seine elektrischen Eigenschaften verleihen Funktionen in den Protonenübertragungsprozessen, die in Mitochondrien stattfinden.
Obwohl Zellen ohne dieses Phospholipid überleben können, haben einige Studien festgestellt, dass sie für die optimale Funktionsweise erforderlich ist.
Verweise
- Harayama, t., & Riezman, h. (2018). Verständnis der Vielfalt der Membranlipidzusammensetzung. Nature Reviews Molekulare Zellbiologie, 19(5), 281-296.
- Luckey, m. (2008). Biologie -Strukturmembran: mit biochemischen und biophysikalischen Grundlagen. Cambridge University Press.
- Murray, r., Bender, d., Botham, k., Kennelly, p., Rodwell, v., & Weil, p. (2009). Harpers illustrierte Biochemie (28. ed.). McGraw-Hill Medical.
- Van Meer, G., Voelker, d. R., & Feigenson, G. W. (2008). Membranlipide: Wo sie sind und wie sie sich verhalten. Nature Reviews, 9, 112-124.
- Vance, j. UND., & Vance, D. UND. (2008). Biochemie von Lipiden, Lipoproteinen und Membranen. In Neues Verständnis Biochemie Vol. 36 (4. Aufl.). Elsevier.
- « Zytosinstruktur, Funktionen, Eigenschaften, Synthese
- Diacilglycerinstruktur, Biosynthese, Funktionen »