

Enolasestruktur, Wirkmechanismus, Funktionen
- 3992
- 865
- Said Ganzmann
Der Enolase Es ist das Enzym, das für die Durchführung der Umwandlung von D-2-Phosphoglycerat (2pga) in Phosphoenolpyruvat (PEP) in Glykolyse und Reverse-Reaktion in der Gluconeogenese, zwei Stoffwechselrouten, die Teil des Zellsenergie-Metabolismus sind.
Die Entscheidung, diese Reaktion in die eine oder andere Richtung zu katalysieren. Das heißt, die Bedürfnisse der Anpassung Ihres Stoffwechsels an die Verschlechterung oder die Synthese, um Energie zu erhalten. Unverzichtbar für die Verwirklichung seiner lebenswichtigen Prozesse.
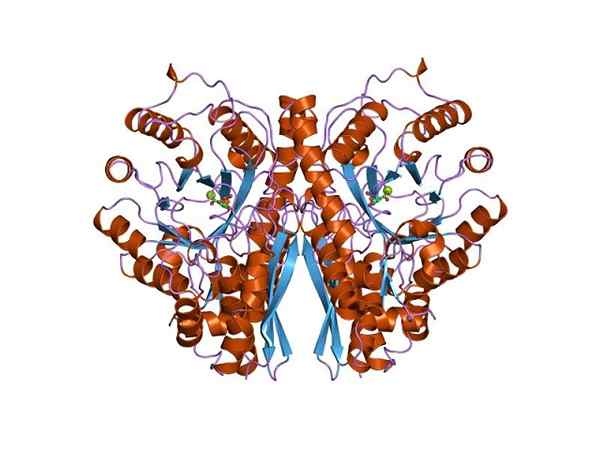
Dreidimensionale Struktur von Enolasa. Von Jawahar Swaminathan und MSD -Mitarbeitern des Europäischen Bioinformatikinstituts [Public Domain (https: // CreativeCommons.Org/lizenzen/by-sa/4.0)] aus Wikimedia Commons. Da beide Stoffwechselwege zum Zentrum des zentralen Stoffwechselbaums der Lebewesen gehören, ist es nicht seltsam, dass die Aminosäuresequenz dieses Proteins in Archae, Bakterien und Eukaryoten erhalten bleibt. Und deshalb zeigen die ähnlichen katalytischen Eigenschaften.
Die Lage der Enolase in der Zelle ist auf Cytosol beschränkt, ein Kompartiment, in dem sowohl die Glykolyse (auch Glykolyse genannt) als auch in den meisten Organismen die Gluconeogenese stattfinden.
Es wurde jedoch auch in anderen Zellkompartimenten wie Plasmamembran vieler Krebszellen und Zellen nachgewiesen. Dort scheint es an der Erleichterung von Zellverbreitungsprozessen beteiligt zu sein, eine völlig andere Funktion als die klassische Funktion.
Enzyme, die in der Lage sind, mehr als eine Funktion auszuführen, wie die Enolase, werden als Mondlicht -Enzyme bezeichnet.
[TOC]
Struktur
Die quartäre Struktur des Linus oder nicht in seinen Liganden wurde in einer großen Anzahl von prokaryotischen und eukaryotischen Individuen bestimmt.
Jedes Monomer präsentiert zwei Domänen: eine kleine Amino-terminale Domäne und eine größere Carboxylterminaldomäne. Die N-terminale Domäne besteht aus drei α- und vier β-Blättern. Während der C-terminale besteht aus acht β-Blättern, die sich zwischen ihnen wechseln und ein β-Barrel bilden, das von acht α-Propellern umgeben ist.
Darüber hinaus werden in jedem Monomer, die als "Konformationsstelle" und "katalytische Stätte" bezeichnet wurden, zwei Gewerkschaftsstellen für zweiwertige Kationen gefunden, die als "Konformationsstelle" und "katalytische Stelle" bezeichnet wurden. Das erste ist nicht sehr selektiv und kann sich in Abwesenheit eines Substrats einer Vielzahl von trennen Kationen anschließen.
Kann Ihnen dienen: Sphingolipids: Was sind, Eigenschaften, Funktionen, SyntheseWährend der zweite bindet an die Ionen, nachdem sich das Substrat dem Enzym angeschlossen hat. Die Vereinigung der Ionen an beide Standorte ist von entscheidender Bedeutung, damit die Reaktion fortgesetzt wird.
Schließlich ist es wichtig zu erwähnen, dass in den Homodimeren die Monomere durch die Aufrechterhaltung einer parallele Orientierung verbunden sind. Daher beschränkt sich das aktive Zentrum auf die von dieser Gewerkschaft gebildete zentrale Region.
Allerdings nimmt nur einer der beiden Monomere an Katalyse teil. Dies erklärt, dass die Fähigkeit von Monomeren, die Reaktion unter experimentellen Bedingungen auszuführen.
Wirkmechanismus
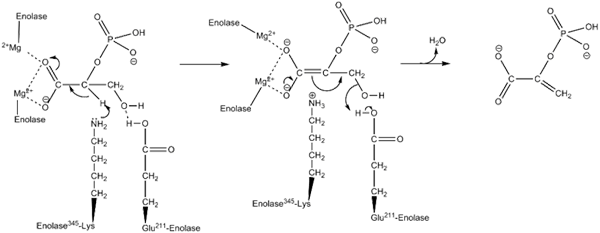 Wirkungsmechanismus, der durch die Enzyminolase verwendet wird. Von kThompson08 in englischer Wikipedia [Public Domain (https: // CreativeCommons.Org/lizenzen/by-sa/4.0)] aus Wikimedia Commons.
Wirkungsmechanismus, der durch die Enzyminolase verwendet wird. Von kThompson08 in englischer Wikipedia [Public Domain (https: // CreativeCommons.Org/lizenzen/by-sa/4.0)] aus Wikimedia Commons. Strukturstudien sowie solche, die die Bestimmung der kinetischen und physikalisch -chemischen Eigenschaften von Enolasa ermöglicht haben, haben es uns ermöglicht, ihren Wirkungsmechanismus zu verstehen.
Die Art und Weise, wie das Enzym die Katalyse der Reaktion durchführt, ist sehr interessant. Obwohl ein einzelnes Substrat beteiligt ist, ist ein geordneter sequentieller Mechanismus derjenige, der vorgeschlagen wurde.
Dies beginnt mit der Vereinigung eines Mg2+ zur Konformationsstelle eines der Monomere. Kontinuierlich mit der Vereinigung des Substrats zum aktiven Zentrum, gefolgt von der Vereinigung eines zweiten Ion. Zu diesem Zeitpunkt bleibt der MG2+ mit der Konformationsstelle verbunden.
In der gleichen Weise, um die Realisierung der Reaktion des durchschnittlichen Enzyms in erster Linie zu begünstigen, die Erzeugung eines Carbanion -Vermittlers. Dies geschieht dies dank der Wirkung eines grundlegenden Aminosäurerests.
Nachfolgend erfolgt eine Carbon -3 -Hydroxylentfernung durch die Wirkung eines sauren Rückstands aus dem Enzym. Zu diesem Zeitpunkt wird die Vereinigung beider Kohlenstoffe mittels einer doppelten Bindung, die PEP bildet, durchgeführt. Auf diese Weise wird die Reaktion gipfelt.
Es kann Ihnen dienen: Monera Kingdom: Merkmale, Klassifizierung und BeispieleFunktionen
Viele der bisher untersuchten Enzyme können eine Vielzahl von Funktionen ausführen, die nicht mit ihrer "klassischen Funktion" in verschiedenen Zellkompartimenten zusammenhängen. Diese Enzyme wurden als "Mondlicht" -Ezyme bezeichnet.
In diesem Sinne kann Enolase als Enzym -Mondlicht betrachtet werden, da zahlreiche Funktionen, die sich ihrer klassischen Funktion widersetzen.
Einige dieser Funktionen sind die folgenden:
- Nehmen Sie sowohl die Aufrechterhaltung der Zellform als auch an vesikulärem Verkehr bei der Interaktion mit Zytoskelett -Proteinen beteiligen.
- Im Kern von Säugetierzellen wirkt ein Transkriptionsfaktor, der die Expression von Genen reguliert. Zusammenarbeit bei der Aufrechterhaltung der Stabilität des RNM im Abbau in Bakterien.
- Bei Krankheitserregern, wie z Streptococcus pneumoniae Und Trypanosoma Cruzi, Es scheint, dass es als wichtiger Virulenzfaktor wirkt.
- Es wurde auch festgestellt, dass in Streptococcus pyogenes, Die Enolase wird in das extrazelluläre Medium ausgeschieden, der die Gewebeverschlechterung und den Umgehung des Wirtssystems erleichtert.
- Auf der Oberfläche von Tumorzellen wird es durch Verbesserung der Metastasierung exprimiert.
Enolase und seine Beziehung zu Zellverbreitungsmechanismen
Zahlreiche Krankheitserreger sowie Tumorzellen exprimieren in ihrer Membran oder vertreiben die extrazellulären Medium -Proteasen, die Proteinproteine abbauen können.
Diese Kapazität ermöglicht es diesen Zellen, das Gewebe durchzubrechen und sich im gesamten Wirtsorganismus schnell auszubreiten. Somit die Umgehung des Immunsystems und damit die Einrichtung der Infektion begünstigen.
Auch wenn Enolasa keine Proteaseaktivität hat, nimmt sie an der Verbreitung vieler Krankheitserreger in seinem Wirt sowie Tumorzellen während der Zielstastase teil.
Dies erreicht es dank der Tatsache, dass es auf der Oberfläche dieser Zellen exprimiert wird, die als Plasminogenrezeptor fungieren. Letzteres ist das Zimogen einer Serinschutz-Protase, das als Plasminat bekannt ist und Teil des fibrinolytischen Systems ist und extrazelluläre Matrixproteine abbaute.
Daher ist die auf der Oberfläche exprimierte Enolase eine Strategie, die diese Zellen erworben haben, um die Infektion zu etablieren und erfolgreich zu verbreiten.
Es kann Ihnen dienen: sympatrische Speziation: Konzept, Merkmale und BeispieleDiese Strategie besteht aus zwei Prozessen:
- Die Ausweichen des Immunsystems des Wirts. Da diese Zellen aus einem Wirtsprotein beschichtet sind, werden sie von Immunsystemzellen ignoriert, die mit Krankheitserregern assoziierte Krankheitserreger erkennen.
- Nach der Aktionsverbreitung von Plasminogen im Plasminat. Deren Teilnahme am Abbau extrazellulärer Matrixproteine und dann die schnelle und wirksame Verbreitung erleichtert.
Verweise
- Avilan L., Gualdron-Lopez M., Quiñones W., González-González L., Hannaert V., Michels PAA, Concepción JL. Enolase: Ein wichtiger Akteur im Stoffwechsel und ein wahrscheinlicher Virulenzfaktor von Trypanosomatiden-Parasiten-Perspektiven für seine Verwendung als therapeutisches Ziel. Enzymforschung. 2011 Vol. Artikel ID932549, 14 Seiten.
- Bhowmick I, Kumar N, Sharma S, Coppens I, Jarori GK, Plasmodium falciparum Enosoase: stadienspezifische Expression und subzelluläre Lokalisation. Malaria Journal. 2009; 8 (1). Artikel 179.
- Tag I, Peshavaria M, Quinn GB, eine differentielle molekulare Uhr in der Enoase -Isoprotein -Evolution. Zeitschrift für Molekulare Evolution. 1993; 36 (6): 599-601.
- der Torre-Scuder E, Manzano-Román R, Pérez-Sánchez R, Siles-Lucas M, Oleaga A. Klonierung und Charakterisierung von plasminogenbindenden Oberflächen -soziierten Enolaase aus Schistosoma Bovis. Tierärztliche Parasitologie. 2010; 173: 73-84.
- Dinovo EC, Boyer PD. Isotopensonden des Enolase -Reaktionsmechanismus. Anfangs- und Äquibium -Isotopen -Wechselkurse: Primär- und Sekunden -Isotopeneffekte. J Biol Chem. 1971; 246 (14): 4586-4593.
- Kaberdin VR, Lin-chao S, entschlüsselt neue Rollen für kleinere Komponenten des E. Coli RNA Abbau. RNA -Biologie. 2009; 6 (4): 402-405.
- Keller A, Peltzer J, Carpentier G. Wechselwirkungen von Enolas. Biochimica et Biophysica Acta. 2007; 1770 (6): 919-926.
- Lung J, Liu KJ, Chang JY, Leu SJ, Shih NY. MBP-1 wird effektiv durch ein alternatives Transkript des Enri1-Gens kodiert, das durch proteasomabhängige Proteinverschiebung reguliert wird. FEBS Journal. 2010; 277 (20): 4308-4321.
- Pancholi v. Multifunktionale α-Enolase: seine Rolle bei Krankheiten. Zell- und molekulare Biowissenschaften. 2001; 58 (7): 902-920.
- Poyner RR, Cleland WW, Reed GH. Rolle von Metall INS in der Katalyse durch Enolase. Ein geordneter kinetischer Mechanismus für ein einzelnes Substratenzym. Biochemie. 2001; 40: 9008-8017.
- Segovia-Gamboa NC, Chávez-Munguía B., Medina-Flores a, Entamoeba Invadens, Enkystationsprozess und Enolase. Experimentelle Parasitologie. 2010; 125 (2): 63-69.
- Tanaka M, Sugisaki K, Nakashima K, Wechsel der translatablen mRNAs für Enoase -Isozyme während der Entwicklung von Chickn -Skelettmuskeln. Biochemische und biophysikalische Forschungskommunikation. 1985; 133 (3): 868-872.