

Sphingolipids Was sind Merkmale, Funktionen, Synthese
- 2971
- 391
- Said Ganzmann
Was sind Spylingolipide?
Der Sphingolipids Sie repräsentieren eine der drei großen Lipidfamilien, die in biologischen Membranen vorhanden sind. Wie Glycephospholipide und Sterole sind sie amphipatische Moleküle mit einer hydrophilen polaren Region und einer hydrophoben apolaren Region.
Sie wurden erstmals 1884 von Johann L beschrieben. W. Thudichum, der drei Sphingolipiden (Sphingomyeline, Gehirn und Gehirnhirn) beschrieb, die zu den drei bekannten Klassen gehören: Phosphosphingolipide, Glykosphingolipide neutral und saure.
Im Gegensatz zu Glyceophospholipiden werden Sphyingolipide nicht auf einem 3-phosphat-Glycerolmolekül als Hauptskelett gebaut, sondern sind Verbindungen, die von Sphiny abgeleitet sind, ein Aminoalkohol mit einer langen Kohlenwasserstoffkette, die von einer Amida-Bindung gebunden ist.
UNDStruktur der Sphingolipide
Alle Sphingolipide stammen aus einem L-Serin, das mit einer langkettigen Fettsäure kondensiert wird, um die Sphingoid-Base zu bilden, die auch als Langkettenbase (LCB) bekannt ist, bildet.
Die häufigsten Grundlagen sind Sphinganin und Sphinxin, die sich in Gegenwart einer Doppelbindung voneinander unterscheiden trans Zwischen den Kohlenstoffen 4 und 5 der Fettsäure des Sphinxin.
Kohlenstoffe 1, 2 und 3 des Sphinxin sind strukturell analog zu Glycerolglycerophospholipiden Glycerin. Wenn Amida -Bindungen eine Fettsäure an Kohlenstoff 2 des Sphin binden.
Langkettige Fettsäuren, die die hydrophoben Regionen dieser Lipide bilden, können sehr vielfältig sein. Die Längen variieren 14 bis 22 Kohlenstoffatome, die unterschiedliche Sättigungsgrade aufweisen können, normalerweise zwischen den Kohlenstoff 4 und 5.
In den Positionen 4 oder 6 können sie Hydroxilli -Gruppen und Doppelbindungen in anderen Positionen oder sogar Auswirkungen wie Methylgruppen haben.
Eigenschaften
Die durch Amida -Bindungen an die Keramiden verbundenen Fettsäureketten sind häufig gesättigt und sind in der Regel größer als die in Glycerophospholipiden, was für die biologische Aktivität von entscheidender Bedeutung zu sein scheint.
Ein charakteristisches Merkmal des Sphingolipid -Skeletts ist, dass sie eine positive Nettobelastung zum neutralen pH -Wert haben können, das bei Lipidmolekülen seltsam ist.
Jedoch der PkA der Aminogruppe ist in Bezug auf ein einfaches Amin zwischen 7 und 8 niedrig, so.
Die traditionelle Klassifizierung von Sphingolipiden ergibt.
Kann Ihnen dienen: Hepadnavirus: Eigenschaften, Morphologie, BehandlungFunktionen
Sphingolipide sind für Tiere, Pflanzen und Pilze sowie in einigen prokaryotischen Organismen und Viren von wesentlicher Bedeutung.
-Strukturfunktionen
Sphingolipide modulieren die physikalischen Eigenschaften von Membranen, einschließlich ihrer Fließfähigkeit, Dicke und Krümmung. Modular Diese Eigenschaften haben ihnen auch einen direkten Einfluss auf die räumliche Organisation von Membranproteinen.
In Lipid "Rafts"
In biologischen Membranen können dynamische Mikrodomänen mit geringerer Flüssigkeit nachgewiesen werden, die durch Cholesterin- und Sphingolipidmoleküle gebildet wird, die als Lipidbalsas bezeichnet werden.
Diese Strukturen treten natürlich auf und halten eine enge Beziehung zu umfassenden Proteinen, Zelloberflächenrezeptoren und Signalproteinen, Transportern und anderen Proteinen mit Glycosylphosphatidylinitol (GPI) -Anker (GPI) (GPI).
-Signalisierungsfunktionen
Sie haben Funktionen wie Signalmoleküle, die als zweite Boten oder als sekretierte Liganden für Zelloberflächenrezeptoren fungieren.
Als sekundäre Boten können sie an der Regulation der Calciumhomöostase, der Zellwachstum, der Tumorogenese und der Unterdrückung der Apoptose teilnehmen. Darüber hinaus hängt die Aktivität vieler integraler und peripherer Membranproteine von ihrer Assoziation mit Sphingolipiden ab.
Viele interzelluläre und Zellwechselwirkungen mit ihrer Umgebung hängen von der Exposition der verschiedenen polaren Gruppen der Sphingolipide in Richtung der äußeren Gesicht der Plasmamembran ab.
Die Vereinigung von Glykosphingolipiden und Lectinas ist für die Assoziation von Myelin mit Axonen, die Adhäsion von Neutrophilen zum Endothel usw. von entscheidender Bedeutung.
Nach Produktprodukten seines Stoffwechsels
Die wichtigsten Signalsphingolipide sind die langkettigen Basen oder die Sphinxe und Ceramiden sowie ihre phosphorylierten Derivate, wie das 1-phosphat-Sphygosin.
Die Metabolismusprodukte vieler Sphingolipiden aktivieren oder hemmen mehrere weiße Downstants (Kinase -Proteine, Phosphatose und andere), die das zelluläre Verhalten so komplex wie Wachstum, Differenzierung und Apoptose kontrollieren.
-Als Membranrezeptoren
Einige Krankheitserreger verwenden Gloysphingolipiden als Rezeptoren, um ihren Eingang zu Wirtszellen zu vermitteln oder Virulenzfaktoren für sie zu liefern.
Es wurde gezeigt, dass trotze Hauptrolle an mehreren Zelleignissen wie Sekretion, Endozytose, Chemiotaxis, Neurotransmission, Angiogenese und Entzündung beteiligt ist.
Sie sind auch am Membranverkehr beteiligt, sodass sie die Internalisierung von Rezeptoren, Ordnung, Bewegung und Fusion sekretorischer Vesikel als Reaktion auf verschiedene Reize beeinflussen.
Sphingolipid -Gruppen
Es gibt drei Sphingolipid -Unterklassen, die alle aus dem Ceramid abgeleitet sind und die sich von polaren Gruppen voneinander unterscheiden, nämlich: Spylingomyeline, Glykolipide und Gangliasiden.
Sphingomyielinas
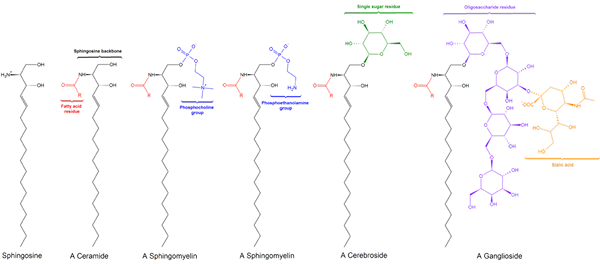
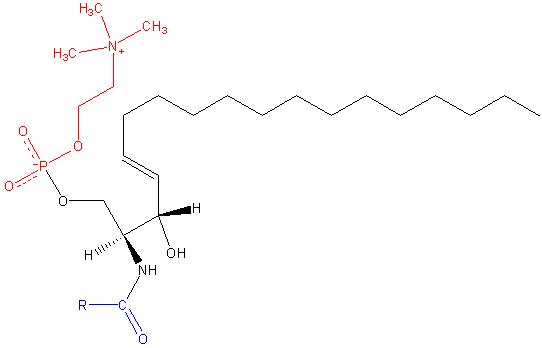 Sphingomilein. Schwarz: Sphinxin. Rot: Phosphocolin. Blau: Fettsäure.
Sphingomilein. Schwarz: Sphinxin. Rot: Phosphocolin. Blau: Fettsäure. Diese enthalten Phosphocolin oder Phosphoethanolamin als polare Kopfgruppe, daher werden sie zusammen mit Glyceophospholipiden als Phospholipide klassifiziert. Sie sehen natürlich Phosphatidylcoline in dreidimensionaler Struktur und allgemeinen Eigenschaften aus, da sie in ihren polaren Köpfen keine Ladung haben.
Kann Ihnen dienen: Enolase: Struktur, Wirkmechanismus, FunktionenSie sind in den Plasmamembranen von Tierzellen vorhanden und sind besonders in Myelin reichlich vorhanden, einer Schote, die die Axone einiger Neuronen umgibt und isoliert.
Glykolipide oder neutrale Glykosphingolipide (ohne Last)
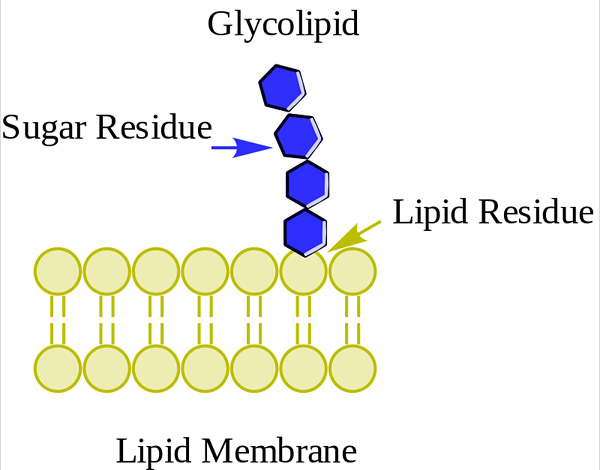 Glucolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativecommons.Org/lizenzen/by-sa/4.0)] aus Wikimedia Commons
Glucolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativecommons.Org/lizenzen/by-sa/4.0)] aus Wikimedia Commons Sie befinden sich hauptsächlich auf der Außenseite der Plasmamembran und haben einen oder mehrere Zucker als polare Kopfgruppe, die direkt mit dem Kohlenstoffhydroxyl 1 des Ceramid -Teils vereint sind. Sie haben keine Phosphatgruppen. Da pH 7 keine Ladung hat, werden sie neutrale Glykolipide bezeichnet.
Cerebroside haben ein einzelnes Zuckermolekül, das mit Ceramid verbunden ist. Diejenigen, die Galactose enthalten. Globoside sind Goustoesphingolipide mit zwei oder mehr Zucker, normalerweise D-Glykose, D-Galactose oder N-Acetyl-D-Galactosamin.
Gangliósidos oder Glykosphingolipide
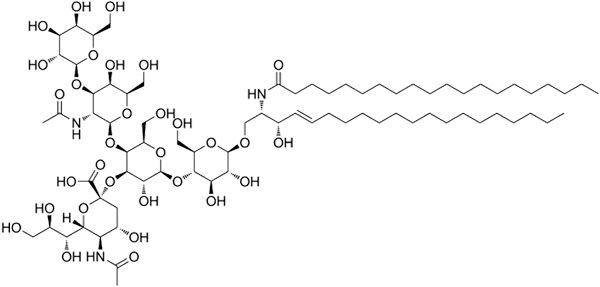 GM1 Ganglosid -Struktur
GM1 Ganglosid -Struktur Dies sind die komplexesten Sphingolipide. Sie haben Oligosaccharide als polare Kopfgruppe und einen oder mehrere terminale N-Acetylmuraminsäure-Reste, auch Siralsäure bezeichnet. Siaminsäure bringt die negative Belastung auf pH 7 in die Ganglien, die sie von neutralen Glykosphingolipiden unterscheidet.
Die Nomenklatur dieser Art von Sphingolipiden hängt von der Menge an Sinalsäureabfällen ab, die im Oligosaccharidabschnitt des Polarkopfes vorhanden sind.
Synthese
Das Long -Chain- oder Sphiny -Basenmolekül wird im endoplasmatischen Retikulum (ER) synthetisiert, und die Zugabe der polaren Gruppe am Kopf dieser Lipide tritt später im Golgi -Komplex auf. Bei Säugetieren kann auch in Mitochondrien eine gewisse Synthese von Sphingolipiden auftreten.
Nach Abschluss ihrer Synthese im Golgi -Komplex werden die Sphingolipide durch Mechanismen, die durch Vesikel vermittelt werden, in andere Zellkompartimente transportiert.
Die Biosynthese der Sphingolipiden besteht aus drei grundlegenden Ereignissen: der Synthese der langkettigen Basen, der Biosynthese der Ceramide durch die Vereinigung einer Fettsäure durch eine Amida -Bindung und schließlich die Bildung der komplexen Sphingolipide mithilfe der Gewerkschaft von polaren Gruppen in Kohlenstoff 1 der Sphingoide -Basis.
Neben der Synthese von novo, Sphingolipiden können sich auch durch Ersatz oder Recycling der langkettigen Basen und Ceramiden bilden, die den Sphingolipid -Pool füttern können.
Ceramid -Skelettsynthese
Die Biosynthese der Ceramid, das Skelett der Sphingolipiden, beginnt mit der Decarboxylerationskondensation eines Palmitail-CoA-Moleküls und eines L-Sernin. Die Reaktion wird durch eine heterodimere, pyridoxale Phosphat-abhängige pyridoxale phosphat-abhängige Serinpalmitailase (SPT) katalysiert und das Produkt ist das 3-Zo-Dihydroesfingosin-Ze.
Kann Ihnen dienen: microbiota: Komposition, Funktionen und wo ist esDieses Enzym wird durch β-Phaal-L-Alanine und L-Cicloserinas gehemmt. In Hefen wird es von zwei Genen codiert, während es bei Säugetieren drei Gene für dieses Enzym gibt. Das aktive Zentrum befindet sich im zytoplasmatischen Gesicht des endoplasmatischen Retikulums.
Die Rolle dieses ersten Enzyms ist in allen untersuchten Organismen erhalten. Es gibt jedoch einige Unterschiede zwischen Taxa, die mit der subzellulären Position des Enzyms zu tun haben.
Das 3-Zoesfinganin wird anschließend durch NADPH-abhängige 3-Zo-Ketosphinganin reduziert. Dihydroceramid-Synthase (Sphinganin N-Acil Transferase) Acetyla dann Sphinanin, um Dihydroceramid zu produzieren. Das Ceramid wird dann durch die Dihydroceramid -Deaturase/Reduktase gebildet, die eine Doppelbindung einführt trans In Position 4-5.
Bei Säugetieren gibt es zahlreiche Ceramid -Isoformen -Syntasas, die jeweils einer spezifischen Fettsäurekette zu den Basen langer Ketten verbinden. Daher bieten Ceramides -Synthesas und andere Enzyme, ENGASAS, die Hauptvielfalt von Fettsäuren in Sphingolipiden an.
Stoffwechsel
Der Abbau der Sphingolipide wird von Glycohydroles und Sphingomyelinase -Enzymen durchgeführt, die für die Entfernung der Modifikationen der polaren Gruppen verantwortlich sind. Andererseits regenerieren die Ceramidasen die langen Kettenbasen aus den Ceramiden.
Die Ganglien werden durch eine Reihe von lysosomalen Enzymen abgebaut, die den Schritt für Schritt der Zuckereinheiten katalysieren und am Ende ein Ceramid erzeugen.
Ein weiterer Verschlechterung ist die Internalisierung von Sphingolipiden in endozytischen Vesikeln, die an die Plasmamembran zurückgeschickt oder zu Lysosomen transportiert werden, wo sie durch spezifische Säurehydrolase abgebaut werden.
Nicht alle langkettigen Basen werden recycelt, der endoplasmatische Rückhalt. Dieser Abbaumechanismus besteht aus Phosphorylierung anstelle der Akilation der LCBs, was zu Signalmolekülen führt.
Verordnung
Der Metabolismus dieser Lipide wird auf mehreren Ebenen reguliert, eines davon ist die der von der Synthese zuständigen Enzyme, ihren post -translationalen Modifikationen und denselben Mechanismen derselben.
Einige Regulationsmechanismen sind zellspezifische Spezifikationen, um entweder die Zeit der Zellentwicklung zu kontrollieren, in der sie produziert werden, oder als Reaktion auf bestimmte Signale.
Verweise
- Bartke, n., & Hannun und. (2009). Bioaktive Spphingolipide: Stoffwechsel und Funktion. Journal of Lipid Research, fünfzig, 19.
- Breslow, d. K. (2013). Sphingolipid -Homöostase im endoplasmatischen Retikulum und darüber hinaus. Cold Spring Harbor Perspektiven in Biologie, 5(4), A013326.