

Muskarinrezeptoren Struktur, Typen und ihre Funktionen, Antagonisten
- 1865
- 478
- Rieke Scheer
Der Muskarinrezeptoren Es sind Moleküle, die Acetylcholin (ACH) vermitteln und sich in der postsynaptischen Membran von Synapsen befinden, in denen der Neurotransmitter freigesetzt wird. Sein Name kommt von seiner Sensibilität gegenüber dem vom Pilz erzeugten Muscarine -Alkaloid Amanita Muscaria.
Im Zentralnervensystem gibt es mehrere neuronale Sätze, deren Axone Acetylcholin freisetzen. Einige von ihnen enden im Gehirn selbst, während die meisten die motorischen Wege für den Skelettmuskel oder die Effektorwege des autonomen Nervensystems für die Drüsen sowie die Herz- und glatten Muskeln ausmachen.
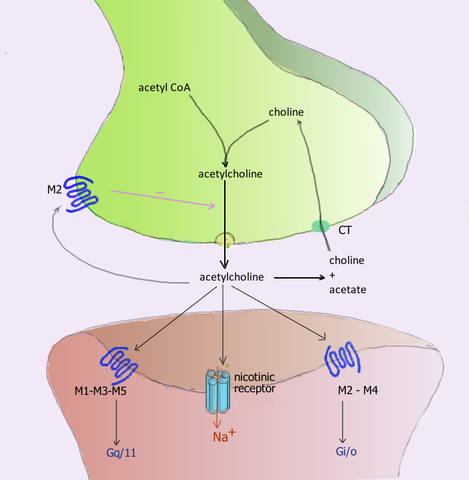
Der Acetylcholin -Neurorezeptor während der Synapse und deren jeweiligen Rezeptoren in der postsynaptischen Membran (Quelle: Benutzer: Pancrat [GFDL 1.2 (http: // www.Gnu.Org/lizenzen/alte Lizenzen/FDL-1.2.html)] über Wikimedia Commons) Acetylcholin, die in den neuromuskulären Gewerkschaften der aktiven cholinergen Rezeptoren des aktiven Skelettmuskels als Nikotinik freigesetzt werden, für seine Empfindlichkeit gegenüber dem Nikotinalkaloid und die auch in den Synapsen Ganglione des autonomen Nervensystems (SNA) gefunden werden.
Die postganglionischen Neuronen der parasympathischen Aufteilung dieses Systems üben ihre Funktionen aus, indem sie Acetylcholin freisetzen.
 Chemische Struktur des Acetylcholin -Neurotransmitters (Quelle: Neurotokeker [Public Domain] über Wikimedia Commons) [TOC]
Chemische Struktur des Acetylcholin -Neurotransmitters (Quelle: Neurotokeker [Public Domain] über Wikimedia Commons) [TOC]
Struktur
Die Muskarinrezeptoren gehören zur Familie der metabotropen Rezeptoren, mit dem die Rezeptoren, die nicht richtig ionische Kanäle sind.
Der Begriff wird verwendet, um sie von ionotropen Rezeptoren zu unterscheiden, die echte ionische Kanäle sind, die sich von direkter Wirkung des Neurotransmitters öffnen oder schließen, wie es bei den Nikotinrezeptoren der Fall ist.
Innerhalb der metabotropen Rezeptoren sind die Muskarinik in der Gruppe, die als gemusster G bekannt ist, enthalten. Sie aktivieren Phospholipase C (PLC).
Muskarinrezeptoren sind lange umfassende Membranproteine; Sie haben sieben Transmarket -Segmente aus Alpha -Propen, die nacheinander die Lipidmembran -Doppelschicht überqueren. Im Inneren sind sie auf dem zytoplasmatischen Gesicht mit dem entsprechenden G-Protein assoziiert, das die Ligand-Empfänger-Wechselwirkung überträgt.
Es kann Ihnen dienen: Embrioblasten: Entwicklung, Funktionen und SchichtenArten von muskarinischen Rezeptoren und deren Funktionen
Mindestens 5 Arten von Muskarinrezeptoren, die unter Verwendung des Buchstabens M bezeichnet werden, gefolgt von einer Zahl, die identifiziert wurde, nämlich: M1, M2, M3, M4 und M5.
M1-, M3- und M5 -Rezeptoren bilden die M1 -Familie und sind durch ihre GQ- oder G11 -Protein -Assoziation gekennzeichnet, während M2- und M4 -Rezeptoren aus der M2 -Familie stammen und mit dem GI -Protein assoziiert sind.
- M1 -Rezeptoren
Sie sind hauptsächlich im Zentralnervensystem, in den exokrinen Drüsen und im Nodon Autonomic System Ganglia gefunden. Sie sind an GQ -Protein gekoppelt, das das Phospholipase -C -Enzym aktiviert, das in das Phosphatidylinosit (PIP2) in Inositol -Tryposphat (IP3) umwandelt, das intrazellulär und Dicalglycerol (DAG) freigibt, das Proteine c aktiviert C.
- M2 -Rezeptoren
Sie werden grundlegend im Herzen gefunden, hauptsächlich in den Zellen des chineurikulären Knoten.
Herzautomatismus
M2 -Rezeptoren wurden im Nodoauricularknoten (SA) des Herzens gründlicher untersucht, wo sich der Automatismus, der regelmäßig die rhythmischen Anregungen erzeugt.
Die Zellen des chineurikulären Knoten. Die Spannung bleibt jedoch nicht in diesem Wert, sondern erleidet eine fortschreitende Depolarisation zu einem Schwellenwert, das ein neues Aktionspotential auslöst.
Diese progressive Depolarisation ist auf spontane Änderungen der Ionenströme (i) zurückzuführen, einschließlich: K+ (IK1) Ausgangsreduzierung, Erscheinung eines Einstiegsstroms von Na+ (FI) und anschließender Eingabe von Ca ++ (ICAT), bis der Schwellenwert erreicht und ein weiterer Ca ++ (iCal), der für das Aktionspotential verantwortlich ist, wird ausgelöst.
Wenn die Ausgabe von K+ (IK1) sehr niedrig ist und die Eingangsströme von Na+ (if) und Ca ++ (ICAT) hoch sind, tritt die Depolarisation schneller auf. Die entgegengesetzten Änderungen in diesen Strömungen niedrigere Frequenz.
Kann Ihnen dienen: CariocineseDurch Noradrenalin (sympathisch) induzierte metabotrope Veränderungen können diese Strömungen verändern. Der AMPC aktiviert direkt die IF -Kanäle, Proteine A (PKA) Phosphoryila und aktiviert die ICAT -Ca ++ - Kanäle, und die β -GI -Proteingruppe aktiviert den Ausgang von k+.
Muskarinaktion M2
Wenn Acetylcholin, die durch die postganglionischen Terminationen der Herzvagalenfasern (parasympathisch) freigesetzt werden.
Die αI -Untereinheit hemmt die Adenylciclase und reduziert die Produktion von AMPC, wodurch die Aktivität der Kanäle für IF und die der PKA reduziert wird. Diese letzte Tatsache reduziert die Phosphorylierung und Aktivität von Ca ++ - Kanälen für ICAT; Das Ergebnis ist eine Verringerung der depolarisierenden Ströme.
Die Gruppe, die durch die βγ -Untereinheiten des GI -Proteins gebildet wird.
Das gemeinsame Ergebnis ist eine Verringerung der spontanen Depolarisation und eine Verringerung der Herzfrequenz.
- M3 -Rezeptoren
 M3 Muskarinrezeptorschema (Quelle: Takuma-Sa [CC0] über Wikimedia Commons)
M3 Muskarinrezeptorschema (Quelle: Takuma-Sa [CC0] über Wikimedia Commons) Sie können im glatten Muskel (Verdauungssystem, Blase, Blutgefäße, Bronchi), in einigen exokrinen Drüsen und im Zentralnervensystem gefunden werden.
GQ -Protein sind ebenfalls gekoppelt und können auf Lungenebene Bronchokonstriktion verursachen, während sie im Gefäßendothelfreisetzungsstickoxid (NO) wirken und eine Vasodilatation verursachen.
- M4- und M5 -Rezeptoren
Diese Rezeptoren sind weniger charakterisiert und untersucht als die vorherigen. Seine Anwesenheit im Zentralnervensystem und in einigen peripheren Geweben wurde berichtet, aber ihre Funktionen sind nicht klar festgelegt.
Kann Ihnen dienen: Tata Box: Eigenschaften und FunktionenAntagonisten
Der universelle Antagonist für diese Rezeptoren ist Atropin, ein aus der Pflanze extrahierter Alkaloid Belladone atropa, Das bindet an sie mit einer hohen Affinität, die ein Kriterium darstellt, um sie von Nikotinrezeptoren zu unterscheiden, die gegenüber diesem Molekül unempfindlich sind.
Es gibt eine große Anzahl anderer antagonistischer Substanzen, die an die verschiedenen Arten von Muskarinrezeptoren mit unterschiedlichen Affinitäten binden. Die Kombination verschiedener Affinitätswerte für einige von ihnen hat genau zur Einbeziehung dieser Rezeptoren in die eine oder andere der beschriebenen Kategorien gedient.
Eine Teilliste anderer Antagonisten umfasste: Pirenzepin, Metotectramin, 4-Damp, Hymbacin, AF-DX 384, Tripitramin, Darifenacin, PD 102807, AQ RA 741, Pfhhsid, MT3 und MT7; Toxine, die in den Giften des grünen und schwarzen Mambas enthalten sind.
M1 -Rezeptoren haben beispielsweise eine hohe Empfindlichkeit gegenüber Pirenzepin; M2 von Tripitramin, Metctramin und Hybacin; der M3 von 4-damp; Die M4 sind sehr mit MT3 -Toxin und auch mit Hybacin verwandt. Die M5 sind dem M3 sehr ähnlich, aber in Bezug auf sie sind sie für hier weniger verwandt. 741.
Verweise
- Ganong WF: Neurotransmitter und Neuromodulatoren, in: Überprüfung der medizinischen Physiologie, 25. ed. New York, McGraw-Hill Education, 2016.
- González JC: Rolle von muskarinischen Rezeptoren bei der Modulation der GABAergen Transmission im Hippocampus. Speicher, um sich für den Abschluss des Arztes zu entscheiden. Autonome Universität von Madrid. 2013.
- Guyton AC, Hall JE: Rythmische Erregung des Herzens, in: Lehrbuch der medizinischen Physiologie , 13. ED; AC Guyton, JE Hall (Hrsg.). Philadelphia, Elsevier Inc., 2016.
- Piper HM: Herzerregung, in: Physiologie des Menschen -Milben Pathophysiologie, 31. Ed; RF Schmidt et al. (Hrsg.). Heidelberg, Springer Medizin Verlag, 2010.
- Schrader J, Gödeche A, Kelm M: Das Hertz, in: Physiologie, 6. ED; R Klinke et al. (Hrsg.). Stuttgart, Georg Thieme Verlag, 2010.
- Siegelbaum SA, Clafam DE, Schwartz JH: Modulation der synaptischen Übertragung: Second Messengers, In: Principles of Neural Science, 5. Aufl.; E Kandel et al. (Hrsg.). New York, McGraw-Hill, 2013.
- « Vorgeschichte der Zellbiologie, welche Studien, Anwendungen und Konzepte
- Wie reproduzieren sich Pflanzen?? Asexuelle und sexuelle Fortpflanzung »